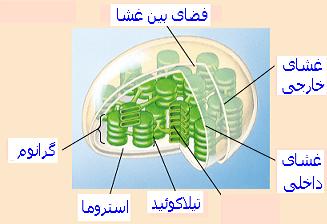
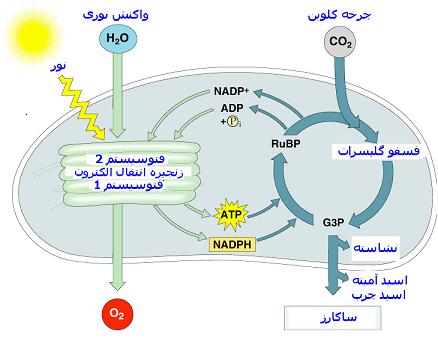
فتوسنتز یکی از فرایندهای حیاتی گیاهان است که غذا و انرژی مورد نیاز گیاهان و سایر موجودات زنده را تامین میکند. این فرایند در دو مرحله انجام میشود. مرحله اول که واکنشهای نوری است. در این مرحله که با استفاده از انرژی نور و حضور آب ، منجر به تولید NADPH و ATP و تصاعد گاز اکسیژن میشوند در دستگاه یا ماشینهای فتوسنتزی به کمک رنگیزههای اصلی و فرعی انجام میگیرند.
واکنشهای نیازمند به نور در گیاهان سبز و جلبکها بوسیله دو سیستم گیرنده نور به نامهای فتوسیستم I و فتوسیستم II انجام میگیرد. بعد از این مرحله واکنشهای بینیاز به نور فتوسنتز انجام میشود که انجام آنها به حضور یا عدم حضور نور وابسته نیست. طی این مرحله با استفاده از انرژی تولید در شده در مرحله نوری فتوسنتز کار تثبیت دیاکسید کربن و تولید قندها انجام میشود.
در هر دوسیستم ، کلروفیلها همراه با رنگیزههای فرعی یک تله گیرندهای را تشکیل میدهند که نور را به دام میاندازد. در سیستم نوری II علاوه بر رنگیزه اصلی P680 رنگیزه فرعی a672 و کلروفیل b و فیکوبیلینها و بعضی از کاروتنوئیدها قرار دارند. سیستم نوری I نیز علاوه بر رنگیزه اصلی P700 دارای رنگیزه فرعی کلروفیل b به مقدار کمتر از سیستم II همچنین رنگیزههای فرعی a مثل a684 نیز هست.
چون این پتانسیل به پتانسیل اکسایش- کاهش سیستم نوری I یا P700 بسیار نزدیک است از این رو الکترونها به آسانی جذب این سیستم میشوند. الکترونها ضمن عبور از زنجیره انتقال الکترون در نقطهای بین پلاستوکینون و سیتوکروم که سقوط یا افت پتانسیل در آنجا زیاد است انرژی خود را از دست میدهند این انرژی مصرف فسفریله کردن ADP و در نتیجه ایجاد ATP در حضور نور (فسفریلاسیون نوری) به مصرف میرسد این فسفریلاسیون با فسفریلاسیونی که در طی فرآیند تنفس صورت میگیرد تفاوت دارد. زیرا مستقل از اکسیژن مولکولی بوده و بدون نیاز به آن در داخل کلروپلاستها رخ میدهد. برای آنکه مولکولهای یونی شده کلروفیل که الکترونهای خود را از دست دادهاند بتوانند کمبود الکترونی را جبران کنند، اجبارا باید الکترون بگیرند.
برای این منظور مولکولهای یونی شده مثبت P680 این کمبود الکترونی را با جذب الکترونهایی که از اکسایش آب آزاد میشوند برطرف میسازند. از اکسایش آب علاوه بر الکترون ، یونهای هیدروژن و هیدروکسید نیز آزاد میشود. که یونهای هیدروکسیل به O2 و H2O تجزیه میشوند و بدین ترتیب اکسیژن فتوسنتزی متصاعد میگردد. یونهای پروتون نیز همراه با الکترونهایی که پس از فعالیت سیستم I به انتهای زنجیره متصل شدهاند صرف احیا NADP و تشکیل NADPH میشوند.
چون در فتوسنتز علاوه بر ATP ، وجود ماده احیا کنندهای جهت تامین هیدروژن یا الکترونها نیز لازم است تا Co2 احیا شده و کربوهیدرات تشکیل شود از این رو فسفریلاسیون نوری یا واکنش تشکیل ATP فتوسنتزی اجبارا با یک واکنش آنزیمی جفت میشود که در کلروپلاستها انجام گرفته و موجب احیای نوکلئوتید پیریدینی NADP میگردد. در این واکنشهای جفت شده یا زوجی نوکلئوتید NADP در حضور نور و آب همراه با ADP و یک مولکول فسفات احیا شده NADPH تبدیل میشود و همزمان با آن ATP نیز شناخته و اکسیژن خارج میشود.
خروج یک مولکول O2 با احیای 2 مول از NADPH و استریفیه شدن 2 مول از فسفات کانی (Pi) همراه است در فتوسنتز باکتریها به جای NADPH نوکلئوتید NADH میسازند.
ضمن بازگشت الکترونها از پلاستوکینون به سیتوکروم F سقوط پتانسیل اکسید و احیا منجر به سنتز ATP میشود و بدین سان هنگام فسفریلاسیون نوری چرخهای ، انرژی نوری به صورت ATP ذخیره میشود بیآنکه احیای CO2 و خروج O2 انجام پذیرد. این نوع فسفریلاسیون توسط طول موجهای بلند ، شدیدتر میشود و این خود موید این است که فقط سیستم I در این فرایند دخالت دارد. به علاوه ترکیباتی که مانع فعالت سیستم II میشوند، برعکس به انجام فرایند فسفوریلاسیون نوری کمک میکنند.
واکنشهای نیازمند به نور در گیاهان سبز و جلبکها بوسیله دو سیستم گیرنده نور به نامهای فتوسیستم I و فتوسیستم II انجام میگیرد. بعد از این مرحله واکنشهای بینیاز به نور فتوسنتز انجام میشود که انجام آنها به حضور یا عدم حضور نور وابسته نیست. طی این مرحله با استفاده از انرژی تولید در شده در مرحله نوری فتوسنتز کار تثبیت دیاکسید کربن و تولید قندها انجام میشود.
 |
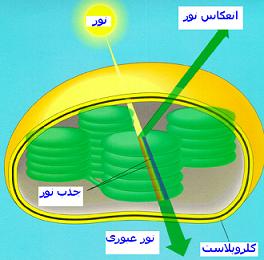
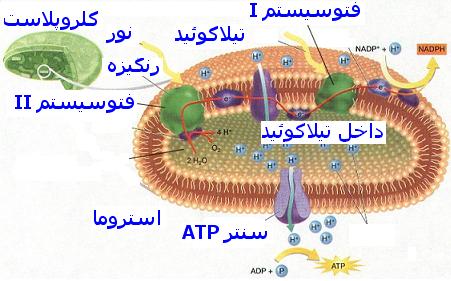
سیستمهای گیرنده نور
برای انجام واکنشهای نوری به همکاری دو گروه مشخص از رنگیزه به نام فتوسیستم (PS) یا سیستم نوری نیاز است. در سیستم نوری I مرکز واکنش یا رنگیزه فعال کلروفیل a است که اوج جذبی آن درطول موج 730 نانومتر است و از این رو P700 نامیده میشود. مرکز واکنش یا رنگیزه فعال سیستم نوری II کلروفیل P680 است که اوج جذبی آن در 682 نانومتر است.در هر دوسیستم ، کلروفیلها همراه با رنگیزههای فرعی یک تله گیرندهای را تشکیل میدهند که نور را به دام میاندازد. در سیستم نوری II علاوه بر رنگیزه اصلی P680 رنگیزه فرعی a672 و کلروفیل b و فیکوبیلینها و بعضی از کاروتنوئیدها قرار دارند. سیستم نوری I نیز علاوه بر رنگیزه اصلی P700 دارای رنگیزه فرعی کلروفیل b به مقدار کمتر از سیستم II همچنین رنگیزههای فرعی a مثل a684 نیز هست.
چگونگی نقل و انتقال الکترون در سیستم نوری II
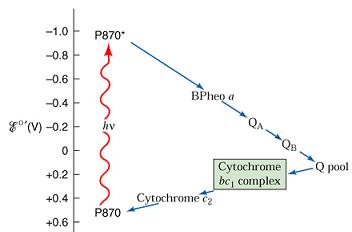
با برخورد فوتونهای نور به برگ گیاه ، ابتدا نخستین تله گیرنده نور یعنی مولکول P680 که در مرکز سیستم نوری II برانگیخته شده، الکترون خود را از دست میدهد و به صورت یونی مثبت درمیآید. این الکترونهای آزاد شده از P680 که انرژی زیادی دارند بلافاصله بوسیله یک سری از مواد انتقال دهنده مانند سیتوکرومها و کینونها که در مجاورت کلروفیل و در غشای تیلاکوئیدی زنجیروار به دنبال هم قرار گرفتهاند منتقل میشود. الکترونهای آزاد شده از مولکول برانگیخته انرژی زیادی دارند و به تدریج با احیا و اکسید شدن مواد ناقل زنجیره الکترون انرژی خود را از دست میدهند و سرانجام به مولکول پلاستوسیانین که پتانسیل اکسایش- کاهش خیلی کمتری دارد، میرسند.چون این پتانسیل به پتانسیل اکسایش- کاهش سیستم نوری I یا P700 بسیار نزدیک است از این رو الکترونها به آسانی جذب این سیستم میشوند. الکترونها ضمن عبور از زنجیره انتقال الکترون در نقطهای بین پلاستوکینون و سیتوکروم که سقوط یا افت پتانسیل در آنجا زیاد است انرژی خود را از دست میدهند این انرژی مصرف فسفریله کردن ADP و در نتیجه ایجاد ATP در حضور نور (فسفریلاسیون نوری) به مصرف میرسد این فسفریلاسیون با فسفریلاسیونی که در طی فرآیند تنفس صورت میگیرد تفاوت دارد. زیرا مستقل از اکسیژن مولکولی بوده و بدون نیاز به آن در داخل کلروپلاستها رخ میدهد. برای آنکه مولکولهای یونی شده کلروفیل که الکترونهای خود را از دست دادهاند بتوانند کمبود الکترونی را جبران کنند، اجبارا باید الکترون بگیرند.
برای این منظور مولکولهای یونی شده مثبت P680 این کمبود الکترونی را با جذب الکترونهایی که از اکسایش آب آزاد میشوند برطرف میسازند. از اکسایش آب علاوه بر الکترون ، یونهای هیدروژن و هیدروکسید نیز آزاد میشود. که یونهای هیدروکسیل به O2 و H2O تجزیه میشوند و بدین ترتیب اکسیژن فتوسنتزی متصاعد میگردد. یونهای پروتون نیز همراه با الکترونهایی که پس از فعالیت سیستم I به انتهای زنجیره متصل شدهاند صرف احیا NADP و تشکیل NADPH میشوند.
 |
چگونگی نقل و انتقال الکترون درسیستم نوری I
در این سیستم مرکز فعال مولکول P700 است که با دریافت الکترونهای منتقل شده از سیستم نوری II برانگیخته میشود و سپس الکترونها را از طریق زنجیرهای از مواد ناقل الکترونی خاص که پتانسیل اکسایش- کاهش خیلی پایینی دارند انتقال میدهد تا به NADP در انتهای زنجیره برسد. الکترونها ابتدا جذب مادهای ناشناخته به نام x میشوند که پتانسیل اکسایش- کاهش ضعیفی دارد و سپس از طریق ناقلین بعدی زنجیره که به ترتیب عبارتند از: فردوکسین ، فلاوپروتئین و NADP منتقل میشوند انتهای این زنجیره NADP بوسیله الکترونهای انتقال یافته و به همراه یونهای پروتون حاصل از تجزیه آب احیا شده و به NADPH تبدیل میشود.فسفریلاسیون نوری
در سال 1954 آرنون و همکارانش نشان دادند که کلروپلاستها آنزیمهای لازم جهت سنتز ATP را دربردارند بطوری که میتوانند در حضور نور ATP بسازند. این ATP بوجود آمده به همراه یک ماده احیا کننده موجب احیا و تثبیت Co2 فتوسنتزی و بالاخره تولید کربوهیدرات در گیاه میشود. آرنون این فرایند ساخته شدن ATP در کلروپلاستها را فسفریلاسیون قتوسنتزی یا فسفریلاسیون نوری نامید.چون در فتوسنتز علاوه بر ATP ، وجود ماده احیا کنندهای جهت تامین هیدروژن یا الکترونها نیز لازم است تا Co2 احیا شده و کربوهیدرات تشکیل شود از این رو فسفریلاسیون نوری یا واکنش تشکیل ATP فتوسنتزی اجبارا با یک واکنش آنزیمی جفت میشود که در کلروپلاستها انجام گرفته و موجب احیای نوکلئوتید پیریدینی NADP میگردد. در این واکنشهای جفت شده یا زوجی نوکلئوتید NADP در حضور نور و آب همراه با ADP و یک مولکول فسفات احیا شده NADPH تبدیل میشود و همزمان با آن ATP نیز شناخته و اکسیژن خارج میشود.
2ADP + 2Pi + 2NADP + 4H2O→2ATP + O2 + 2NADPH + 2H2O
خروج یک مولکول O2 با احیای 2 مول از NADPH و استریفیه شدن 2 مول از فسفات کانی (Pi) همراه است در فتوسنتز باکتریها به جای NADPH نوکلئوتید NADH میسازند.
فسفریلاسیون نوری غیر چرخهای
هنگامی که دو سیستم نوری II , I همزمان با هم و با دخالت آب همکاری میکنند انتقال الکترونهای پر انرژی آزاد شده از کلروفیل برانگیخته توسط فوتونهای نور که با تشکیل NADPH , ATP همراه است مسیری غیر چرخهای را به شکل حرف Z طی میکنند به نحوی که الکترونها پس از عبور از زنجیره انتقال الکترون دیگر به مولکول کلروفیل باز نمیگردند و کمبود یا خلا الکترونی از تجزیه آب جبران میشود به این فرآیند انتقال غیر چرخهای الکترونها که بر اثر همکاری هر دو سیستم II,I صورت میگیرد و به ساخته شدن NADPH , ATP میانجامد فسفریلاسیون نوری غیر چرخهای نیز میگویند.فسفریلاسیون نوری چرخهای
در این فسفریلاسیون که بدون دخالت سیستم II و تصاعد اکسیژن انجام میگیرد فقط سیستم نوری I برانگیخته میشود و الکترونهای برانگیخته از کلروفیل P700 پس از عبور از زنجیره انتقال الکترون همین سیستم با مسییری دایره وارد چرخه دوباره به کلروفیل P700 برمیگردند. و ضمن این بازگشت انرژی خود را از دست میدهند که صرف ساختن ATP میشود.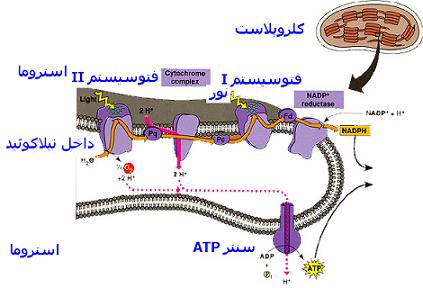 |
علت فرآیند فسفریلاسیون نور چرخهای
فسفریلاسیون نوری چرخهای هنگامی انجام میگیرد که واکنشهای مرحله نوری به دلایلی نظیر نرخ پایین CO2 ، عدم خروج فرآوردههای نهایی فتوسنتز از یاختههای فتوسنتز کننده و در نتیجه عدم مصرف NADPH و بالاخره کافی نبودن ATP حاصل از فسفریلاسیون غیر چرخهای متوقف شوند در چنین مواردی الکترونها پس از احیای فردوکسین بوسیله NADPH گرفته نمیشوند بلکه با دخالت سیتوکروم b به پلاستوکینون و پس به سیتوکروم F و پلاستوسیانین انتقال مییابند و از طریق این مواد مجددا به کلروفیل P700 در سیستم I برمیگردند.ضمن بازگشت الکترونها از پلاستوکینون به سیتوکروم F سقوط پتانسیل اکسید و احیا منجر به سنتز ATP میشود و بدین سان هنگام فسفریلاسیون نوری چرخهای ، انرژی نوری به صورت ATP ذخیره میشود بیآنکه احیای CO2 و خروج O2 انجام پذیرد. این نوع فسفریلاسیون توسط طول موجهای بلند ، شدیدتر میشود و این خود موید این است که فقط سیستم I در این فرایند دخالت دارد. به علاوه ترکیباتی که مانع فعالت سیستم II میشوند، برعکس به انجام فرایند فسفوریلاسیون نوری کمک میکنند.